L'effet bénéfique de la lumière rouge
pour l'oeil
LLLT : Low-Level Light Therapy
Résumé de la méta-étude du Dr Rojas
Departments of Psychology, Pharmacology and Toxicology, University of Texas at Austin, USA.
La lumière
La lumière est une forme d'énergie appelée « énergie lumineuse ». Une onde de radiation électromagnétique unidirectionnelle et caractérisée par la longueur d'onde (la distance entre les « pics » ou les « vallées » de l'onde), la fréquence (le nombre d'oscillations par seconde) et l'amplitude (la différence entre les « Pics » et les « vallées »). Un ensemble complexe d'ondes de différentes amplitudes, fréquences et longueurs d'onde compose la lumière qui se réfléchit sur les objets.
La lumière d'une seule longueur d'onde est appelée monochromatique. En physique quantique, la radiation électromagnétique consiste en photons qui sont des particules d'énergie voyageant à 300.000 km/seconde. La luminosité de la lumière est le nombre de photons et la couleur de la lumière est l'énergie contenue dans chaque Photon. LLLT (Low level Light therapy), La thérapie par de faible niveau de lumière (LLLT = « low level light therapy »), peut être définie par l'usage d'une lumière monochromatique de laser ou de diodes notamment dans le rouge ou le proche infrarouge pour moduler une fonction biologique sans effet thermique.
L'effet de telle action implique la conversion de l'énergie lumineuse en énergie métabolique avec pour effet la modulation du fonctionnement biologique des cellules. On parle de « PHOTOBIOMODULATION » ou « PHOTONEUROMODULATION » quand les cellules nerveuses en sont la cible.
Laser ou diode électroluminescente
La différence avec les lasers est la haute énergie délivrée qui résulte d'un réchauffement des tissus jusqu'à leur destruction lors de dissection, ablation, coagulation ou vaporisation.
C'est la raison pour laquelle on parle au sujet des diodes électroluminescentes (LED) de « LOW LEVEL » (LLLT). L'énergie délivrée via ces diodes est inversement proportionnelle à sa longueur d'onde. L'énergie délivrée par LLLT est trop faible pour causer de la chaleur ou de la destruction par contre, elle l'est assez pour moduler les fonctions cellulaires. La longueur d'onde du rouge ou proche de l'infrarouge a montré son efficacité optimale en induisant des effets bénéfiques aux cellules qui n'apparaissent pas comme spécialisées en photo-pigments.
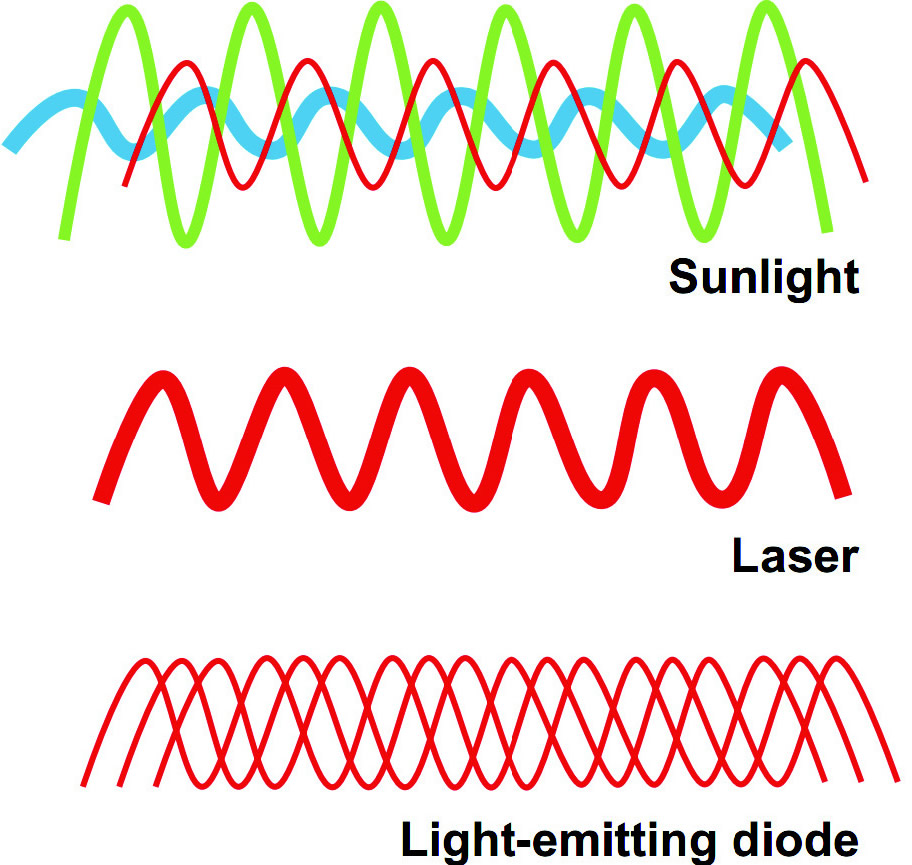
Figure 1 – La lumière du soleil se compose d'une combinaison d'ondes non cohérentes avec des longueurs d'onde couvrant la totalité du spectre visible. En revanche, les lasers émettent une longueur d'onde unique (monochrome). Cette cohérence de longueur d'onde élevée permet la transmission de l'énergie à une densité de puissance élevée. Enfin, la lumière à faible niveau se compose d'ondes monochromatiques ou quasi-monochromatiques en prenant des chemins différents menant à un point cible commun. Les auteurs introduisent la possibilité que la non-cohérence peut être avantageuse à des fins neuro-métaboliques et qu'elle permet l'exposition du tissu nerveux à des longueurs d'onde «thérapeutiques», à des densités de puissance relativement faible pendant le temps nécessaire pour moduler le métabolisme neuronal.
A la différence de la lumière solaire non cohérente et multidirectionnelle, la lumière monochromatique cible la bio-photo modulation (voir Figure 1).
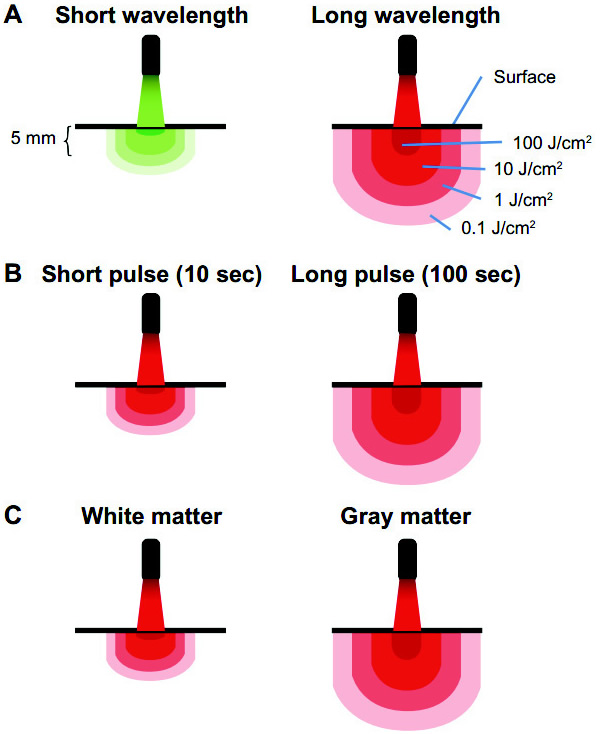 Figure 2 – Principes des interactions lumière-tissu. (A) La lumière aux longueurs d'onde courtes a une pénétration tissulaire faible. La lumière à haute longueurs d'onde affiche une pénétration tissulaire élevée et offre des niveaux d'énergie thérapeutiques à des structures plus profondes. (B) Parce que l'exposition énergétique (J/cm²) est le produit de l'éclairement énergétique (w/cm²) et le temps, l'énergie délivrée aux tissus à la suite d'un éclairement constant peut être augmentée en augmentant le temps d'exposition. Ainsi, la pénétration du tissu peut également être affectée par la durée d'exposition. Lorsque les sources de thérapie par la lumière à faible niveau (LLLT) sont utilisées avec des temps d'exposition élevés, les structures profondes peuvent être traitées avec des quantités d'énergie bio-modulatoire, tout en évitant les effets d'ablation. (C) Enfin, les tissus varient dans leur teneur en photo-accepteur, de transmission, et de temps de relaxation. Cela représente des variations interspécifiques et interrégionales pour la pénétration de la lumière (par exemple, la matière grise contre la substance blanche dans le cerveau).
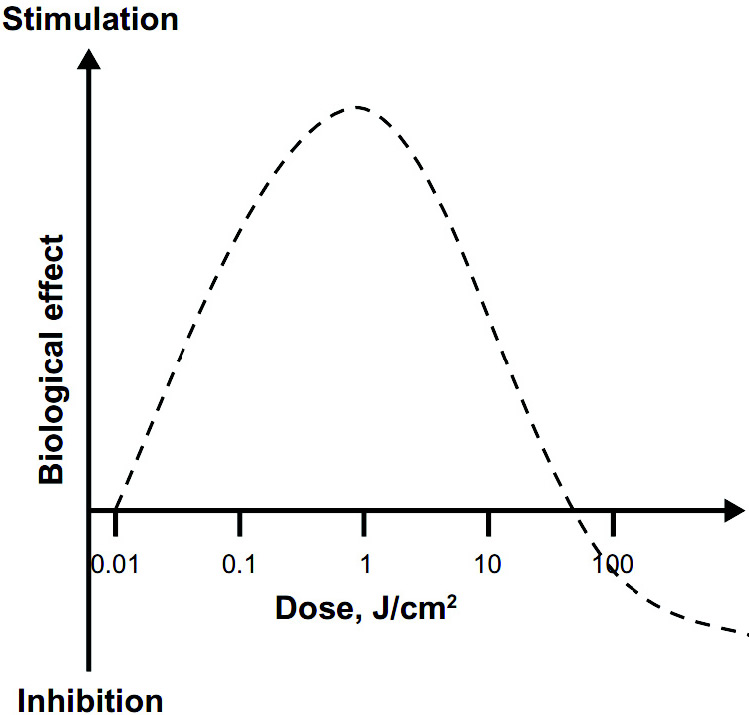
Figure 3 – Les effets hormétiques de la thérapie par la lumière à faible niveau (LLLT) : Les effets LLLT sont caractérisés par des courbes dose-réponse en forme de U inversé, dont les réponses linéaires peuvent être vues seulement à des doses très faibles. Alors que les effets linéaires peuvent être négligeables, les effets stimulants maximaux sont généralement observés à des doses intermédiaires. Une observation clé concernant les effets modulateurs de la lumière dans les tissus est que les réponses maximales, à des doses intermédiaires, ont tendance à représenter moins de deux fois l'augmentation des variables biologiques par rapport aux conditions de référence.
« Photo récepteurs » et « Photo accepteurs »
Il a été montré que les systèmes biologiques contiennent à la base deux types de molécules qui peuvent absorber la lumière : des molécules spécialisées et des molécules non spécialisées.
Des molécules hautement efficaces avec un degré de spécification pour la conversion d'énergie sont connues comme les « photo récepteurs ». Ceci inclus les molécules photo pigment, comme la chlorophylle qui est importante dans la photosynthèse des plantes. Les cônes et les bâtonnets sont quant à eux les « photo récepteurs » de la rétine déterminants pour la fonction visuelle. La melanopsine trouvée dans les cellules ganglionnaires de la rétine est, quant à elle, responsable de la régulation de l'horloge biologique.
D'un autre côté, des molécules non spécialisées qui peuvent absorber la lumière mais qui ne sont pas intégrées à un organe récepteur de lumière sont appelée « photo accepteurs ». Quoique les « photo accepteurs » peuvent potentiellement absorber la lumière, ils font partie habituellement de voies métaboliques qui ne sont pas directement liées avec le traitement de la lumière. Les « photo accepteurs » sont plus abondants et omniprésents que les « photo-récepteurs». Les molécules « photo accepteurs » qui sont capables d'absorber la lumière et d'exercer des changements dans la fonction ou la structure de la cellule se trouvent dans pratiquement toutes les cellules vivantes et pas seulement celles des organes sensoriels spécialisés comme l'œil.
Le « chromophore » est un autre concept en photobiologie. Le « Chromophore » est un fragment particulier dans une molécule « photo-accepteur » ou « photo récepteur » qui est responsable de l'absorption de la lumière.
Habituellement ce sont des « cofacteurs » ou des « ions métal » dans une structure de protéine qui contiennent des électrons qui peuvent passer du stade basal au stade excité (voir loi Boltzmann).
L'excitation induit une changement dans la conformation moléculaire et le métabolisme intracellulaire. L'association entre les chromophores et les enzymes respiratoires a un rôle majeur dans la régulation de la fonction neurale.
Le flux d'énergie électromagnétique se transforme ainsi en flux d'énergie chimique. Une propriété particulière des chromophores est d'absorber des longueurs d'ondes particulières et d'en refléter d'autres. Ainsi l'anneau porphyrine de la chlorophylle a un pic d'absorption à 430 nm et 662 nm. De manière similaire certains cônes sont sensibles à 430 nm (bleu), 530 nm(vert) et 650 nm (rouge) ce qui permet au cerveau une sensibilité relative aux couleurs.
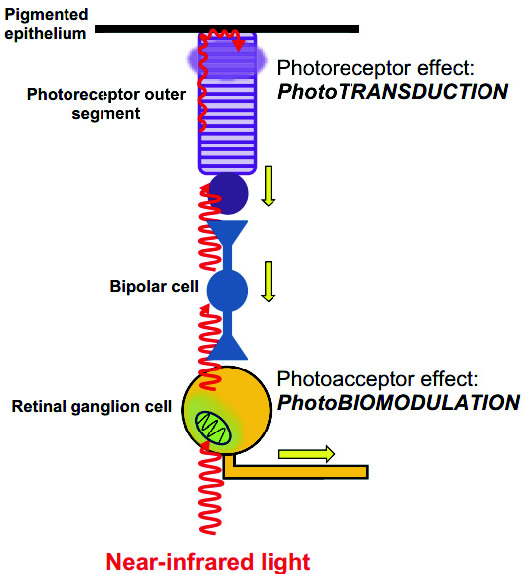 Figure 4 - Les effets différentiels de la lumière sur des photorécepteurs et photo-accepteurs dans la rétine. La lumière atteint la rétine et se déplace à travers les différentes couches de la rétine pour atteindre la couche la plus externe du photorécepteur. Il excite alors le photorécepteur ‘rhodopsine' dans les bâtonnets et les cônes, déclenchant le processus de photo transduction. La photo transduction provoque l'hyperpolarisation des cellules photo réceptrices, des changements dans la neurotransmission et des potentiels d'action (flèches jaunes) dans les cellules bipolaires et les cellules ganglionnaires. Ces effets représentent le début du traitement de l'information visuelle. La lumière peut également exciter directement les photo-accepteurs dans les neurones, y compris des cellules ganglionnaires de la rétine. Le principal photo-accepteur dans le spectre du rouge au proche infrarouge est l'enzyme respiratoire mitochondriale cytochrome C oxydase. Les effets de la lumière sur les cytochrome C oxydase des neurones induisent une modulation des mécanismes bioénergétiques cellulaires qui sont indépendants du traitements visuels. Il est bien connu aujourd'hui que la photo réception à la lumière à d'importants effets biologiques indépendamment des fonctions visuelles. Les cellules ganglionnaire de la rétine, par exemple, fournissent l'information sur la luminance de l'environnement au système de régulation des rythmes circadiens et règle par ailleurs le réflex pupillaire. La mélanopsine absorbe la lumière via un chromophore rétine-aldéhyde ce qui conduit à une modification du potentiel membranaire via une cascade de signalisation avec des G-protéines. La glande pinéale quant à elle contient la pinopsin, un photorécepteur similaire qui est sensible à 470 nm qui est exclusivement exprimé dans cet organe et non exprimé dans la rétine ou ailleurs dans le cerveau. C'est grâce à la pinopsin que le pacemaker circadien contrôle la production de mélatonine indépendamment des voies visuelles (cônes et bâtonnets).
Il est bien connu aujourd'hui que la photo réception à la lumière à d'importants effets biologiques indépendamment des fonctions visuelles. Les cellules ganglionnaire de la rétine, par exemple, fournissent l'information sur la luminance de l'environnement au système de régulation des rythmes circadiens et règle par ailleurs le réflex pupillaire. La mélanopsine absorbe la lumière via un chromophore rétine-aldéhyde ce qui conduit à une modification du potentiel membranaire via une cascade de signalisation avec des G-protéines. La glande pinéale quant à elle contient la pinopsin, un photorécepteur similaire qui est sensible à 470 nm qui est exclusivement exprimé dans cet organe et non exprimé dans la rétine ou ailleurs dans le cerveau. C'est grâce à la pinopsin que le pacemaker circadien contrôle la production de mélatonine indépendamment des voies visuelles (cônes et bâtonnets).
Ciblage moléculaire du traitement par la lumière LLLT
Différentes études ont montré de remarquables effets in vitro sur les cellules vivantes notamment l'accélération du métabolisme des fibroblastes, de la synthèse du DNA et du taux de croissance mais aussi sur la résolution de l'inflammation et sur les processus de guérison.
Le rouge et le proche infra-rouge semblent avoir le meilleur taux de réponse photo-biologique. A ce propos, l'hémoglobine semble remporter la palme des « photoaccepteurs » les plus sensibles dans des tissus aussi variés que les cellules neurales, les fibroblastes et les cellules de l'épithélium. Présents dans les globules rouges, on soupçonne d'autres « photoaccepteurs » intermédiaires avec l'hémoglobine notamment la myoglobine et le cytochrome-C oxydase mais aussi le cytochrome-B oxydase, la superoxyde dismutase, l'oxyde nitrique synthase, la guanylase cyclase et les cryptochromes. Ceci se complique par le fait que la correspondance entre une simple longueur d'onde et une cible moléculaire unique n'est pas directe.
Par exemple, les flavoprotéines, NADH déshydrogénase, ont été identifié comme « photoacceptateurs » à la fois dans le bleu violet et dans le spectre rouge du proche infra-rouge. De même, les terminal oxydases et l'enzyme antioxydant superoxyde dismutase montre aussi un pic d'absorption dans les hautes longueurs d'ondes de 670 nm à 680 nm. On observe de manière générale que l'illumination a un effet sur les mitochondries et augmente la synthèse de l'ATP, de l'acide ribonucleique (ARN), de la synthèse protéique et la consommation d'oxygène (24) et (25). Ceci suggère que les molécules qui sont sensibles à certaines des longueurs d'ondes de la lumière sont des composants de la chaîne respiratoire responsable de la production d'énergie pour les cellules.
De même, les terminal oxydases et l'enzyme antioxydant superoxyde dismutase montre aussi un pic d'absorption dans les hautes longueurs d'ondes de 670 nm à 680 nm.
On observe de manière générale que l'illumination a un effet sur les mitochondries et augmente la synthèse de l'ATP, de l'acide ribonucleique (ARN), de la synthèse protéique et la consommation d'oxygène (24) et (25).
Ceci suggère que les molécules qui sont sensibles à certaines des longueurs d'ondes de la lumière sont des composants de la chaîne respiratoire responsable de la production d'énergie pour les cellules.
L'enzyme cytochrome C Oxydase est aujourd'hui reconnue comme l'enzyme clé ayant un effet photo bioénergétique pour les cellules, spécialement pour les cellules nerveuses de la rétine et du cerveau (28). Cet enzyme jouerait donc le rôle d'appareil de transfert d'énergie.Cette molécule très complexe est couplée étroitement à la demande énergétique, le métabolisme des radicaux libres, le nettoyage cellulaire et l'activation glutaminergique. (30 et 31). Les pics d'absorption de cet enzyme sont 620 nm, 680 nm, 760 nm et 825 nm.
Mécanismes d'actions
Les neurones dépendent de la cytochrome-C Oxydase pour produire leur métabolisme énergétique. Les effets de LLLT ont été subdivisé en deux parties: les effets durant l'exposition à la lumière et les effets après exposition à la lumière. Les effets primaires sont des changements redox des composants de la chaine respiratoires, la production d'ATP et la génération de radicaux libres. Les effets secondaires sont une cascade de réactions biochimiques qui modifient l'homéostasie cellulaire, la neurotransmission, l'expression de gènes et des fonctions enzymatiques. Dans ce contexte, la mitochondrie enverrait des signaux au noyau qui à son tour agirait dans l'expression de gènes adaptatif.
Effet de l'LLT sur le tissu nerveux
Les neurones sont plus que tout autres cellules sujet au métabolisme aérobie. Le traitement par LLLT et son influence sur la chaîne respiratoire des mitochondries peut donc avoir un rôle à ce niveau tant pour les tissus neuraux endommagés que pour les tissus sains en demande énergétique comme le travail intellectuel intense. De multiples études ont prouvé un effet réparateur et régénérateur des composants des cellules nerveuses. (8, 9, 38, 39, 42, 45, 47, 48)
Différentes maladies des yeux peuvent être prises en charge par la LLLT : dégénération rétinienne, dégénérescence maculaire, neuropathie optique, glaucome, Alzheimer, atrophie du nerf optique dont la plupart sont souvent liées à des dysfonctions mitochondriales y compris l'épilepsie myoclonique, l'AVC, etc. La LLLT peut ainsi être globalement considérée comme une thérapie contre les dommages rétiniens, celle-ci contrecarrant les conséquences immédiates des défaillances mitochondriales et étant donc un facteur actif de réparation neurale.
En ce qui concerne les AVC et traumas neurologique la LLLT en transcranial augmente le flux cérébral (86) de 30% avec 1,6 W/cm2.
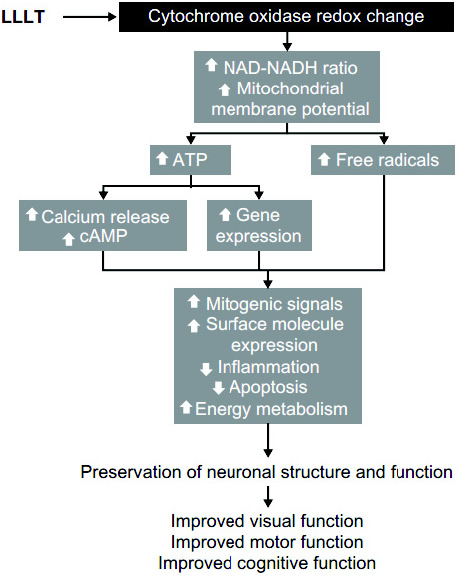 Figure 5 - Les mécanismes intracellulaires de l'action de la luminothérapie à faible niveau (LLLT). Résultats de la photo-bio modulation dans une cascade d'effets pléiotropes intracellulaires. La lumière est absorbée par les chromophores dans le cytochrome oxydase et induit des changements dans son état d'oxydo-réduction. La réaction d'oxydoréduction d'enzymes dans la membrane mitochondriale interne induit notamment un écoulement accéléré d'électrons et des modifications sur le NADH et le potentiel de la membrane mitochondriale. Ces changements facilitent la synthèse de l'adénosine triphosphate (ATP)la molécule d'énergie de la cellule et augmentent la production de radicaux libres. L'augmentation de la disponibilité de l'ATP permet l'activation de kinases qui induisent la libération de calcium et la formation de l'adénosine mono phosphate cyclique (AMPc). Le calcium, l'AMPc, et les radicaux libres agissent comme messagers secondaires et sont capables d'activer différentes voies métaboliques au niveau nucléaire. Il s'ensuit une amélioration de la physiologie neuronale qui se traduit par une amélioration clinique.
L'étude complète
>> Lire l'étude complète en anglais (fichier PDF)
TABLE 1 - Paramètres majeurs de la thérapie LLLT
(low level light therapy)
Références
8. Eells JT, Wong-Riley MT, VerHoeve J, et al. Mitochondrial signal transduction in accelerated wound and retinal healing by near-infrared light therapy. Mitochondrion. 2004;4(5–6):559–567.
9. Wong-Riley MT, Liang HL, Eells JT, et al. Photobiomodulation directly benefits primary neurons functionally inactivated by toxins: role of cytochrome c oxidase. J Biol Chem. 2005;280(6):4761–4771.24. Karu T. Mechanisms of low-power laser light action on cellular level. Proc SPIE. 2000;4159:1–17.
24. Karu T. Mechanisms of low-power laser light action on cellular level. Proc SPIE. 2000;4159:1–17.
25. Karu T. Laser biostimulation: a photobiological phenomenon. J Photochem Photobiol B. 1989;3(4):638–640.
28. Wong-Riley M. Energy metabolism of the visual system. Eye and Brain. 2010;2:99–116.
30. Sakata JT, Crews D, Gonzalez-Lima F. Behavioral correlates of differences in neural metabolic capacity. Brain Res Brain Res Rev. 2005;48(1):1–15.
31. Wong-Riley MT. Cytochrome oxidase: an endogenous metabolic marker for neuronal activity. Trends Neurosci. 1989;12(3):94–101.
38. Liang HL, Whelan HT, Eells JT, et al. Photobiomodulation partially rescues visual cortical neurons from cyanide-induced apoptosis. Neuroscience.
39. Byrnes KR, Wu X, Waynant RW, Ilev IK, Anders JJ. Low power laser irradiation alters gene expression of olfactory ensheathing cells in vitro. Lasers Surg Med. 2005;37(2):161–171.
42. Byrnes KR, Waynant RW, Ilev IK, et al. Light promotes regeneration and functional recovery and alters the immune response after spinal cord injury. Lasers Surg Med. 2005;36(3):171–185.
45. Wollman Y, Rochkind S. In vitro cellular processes sprouting in cortex microexplants of adult rat brains induced by low power laser irradiation. Neurol Res. 1998;20(5):470–472.
47. Rochkind S, Vogler I, Barr-Nea L. Spinal cord response to laser treatment of injured peripheral nerve. Spine (Phila Pa 1976). 1990;15(1):6–10.
48. Anders JJ, Borke RC, Woolery SK, Van de Merwe WP. Low power laser irradiation alters the rate of regeneration of the rat facial nerve. Lasers Surg Med. 1993;13(1):72–82.
86. Uozumi Y, Nawashiro H, Sato S, Kawauchi S, Shima K, Kikuchi M. Targeted increase in cerebral blood flow by transcranial near-infrared laser irradiation. Lasers Surg Med. 2010;42(6):566–576. |